|
Abstract
Keloids are skin abnormalities that are characterized by
excessive deposition of collagen bundles in the dermis.
Patients with keloids complain not only about their cosmetic
appearances, but also about continuous itching and/or
tenderness associated with chronic inflammation. Degradation
of extracellular matrix (ECM) may be upregulated associated
with the expansion of keloids into circumferential skin,
and high metabolic activity of keloid tissues may be
due to increased matrix metalloproteinases (MMPs) activity.
Based on these hypotheses, we examined differences in
expressions of MMP-1, MMP-8, and MMP-13 between keloid-derived
fibroblasts and normal dermal fibroblasts. Since retinoids
are potent inhibitors of MMPs in the treatment of photoaged
skin and cancers, we also examined whether or not tretinoin
affects MMPs expressions of keloid-derived fibroblasts.
The results of real-time PCR and ELISA demonstrated a significant
upregulation of MMP-13 as well as significant downregulation
of MMP-1 and MMP-8 in keloid-derived fibroblasts, both at
mRNA and protein levels. MMP-1 mRNA expression in the control
group was significantly upregulated after the addition of
tretinoin, whereas no significant change was observed in
the keloid group. MMP-8 mRNA expression in the control group
was significantly upregulated with the peak at 12 hours by
tretinoin, while no significant change was observed in the
keloid-derived fibroblasts. In contrast, the remarkably elevated
MMP-13 mRNA expression in the keloid group was significantly
suppressed with the peak suppression at 12 hours after addition
of tretinoin, while MMP-13 mRNA expression in the control
group was not significantly changed.
The decrease in MMP-1 and MMP-8 may contribute to accumulation
of type I and type III collagen in keloid tissues, and this
mechanism may be modulated by molecular interaction with
MMP-13. Tretinoin appeared to reverse the abnormal expression
profile of MMPs in keloid-derived fibroblasts, such as markedly
elevated expression of MMP-13, partly through inactivation
of AP-1 pathway. The present results suggested that tretinoin
may be clinically useful to improve chronic inflammation
seen in keloids and prevent expansion of keloid tissues into
circumferential normal skin.
Introduction
Keloids are skin abnormalities that are characterized by
excessive deposition of collagen bundles in the dermis. In
keloids, the normal wound healing process is derailed from
the normal, resulting in impairment of the balance bewteen
production and degradation of extracellular matrix (ECM),
such as collagens (1). Since fibroblasts play a leading part
in production of ECMs, it is thought that there is a difference
in the cellular function between keloid-derived fibroblasts
and normal ones. However, accumulating data have shown that
there is no significant difference in culture growth, cell
size, population density, and karyotype between these (2).
During normal wound repair, type III collagen appears at
day 2 to 3, followed by type I collagen at day 6 to 7 (3).
The total amount of type I and III collagen increases over
time, whereas the proportion of type III collagen decreases
from 60 % at 1 week after wounding, to 28 % in the mature
scar (4). In keloids, however, the relative amount of type
III collagen remains high compared to normal scars or normal
skin (5). The ratio of α-1(I)-procollagen (a precursor of
type I collagen) mRNA to α-1(III)-procollagen (a precursor
of type III collagen) mRNA is markedly elevated in keloid-derived
fibroblasts compared to that in normal-tissue-derived fibroblasts
in vitro (6). The same tendency is also observed in keloid
tissues in vivo (7). There seems to be a discrepancy between
excessive accumulation of type III collagen in keloid tissues
and an elevated mRNA level of type I procollagen in biosynthesis,
which remains unknown. This discrepancy is explicable if
cytologic aberrations occur at the level of the degradation
of collagen fibers, especially type III collagen. However,
past studies have only shown a normal (8), decreased (9),
increased (10) collagenase activity (more accurately, degradation
activity of type I collagen), and no studies have demonstrated
altered expression of each type of collagenase in keloid
tissues or keloid-derived fibroblasts.
Currently, collagenases are categorized into groups of endopeptidases
with a divalent Zn2+ at the active site involved in ECM remodeling,
matrix metalloproteinases (MMPs). MMP-1 (also known as interstitial
collagenase or collagenase 1), MMP-8 (neutrophil collagenase,
collagenase 2), and MMP-13 (collagenase 3), are the only
mammalian enzymes recognized for their unique ability to
cleave the triple helical domain of fibrillar collagen types
I, II, and III (11). However, each collagenase differs in
the extent to which it cleaves these fibrillar collagen subtypes
in vitro. MMP-1 preferentially degrades type III collagen,
whereas MMP-8 has its greatest activity on type I collagen
(12). Neither MMP-1 nor MMP-8 appears to have any significant
activity against type II and IV collagen. MMP-13 is the most
recently discovered human collagenase, which can degrade
all fibrillar collagen subtypes with almost equal efficacy,
and is the only collagenase with significant activity against
type II and IV collagen (13).
Before establishing novel classification of collagenases
as descrived above, fibroplastic lesions due to deposition
of ECM such as collagen fibers had been classified into two
groups; increased-level group and decreased-level group,
according to their collagenase activities. Rheumatoid arthritis,
osteoarthritis, periodontal diseases, otitis media cholesteatoma,
and malignant tumors belong to the former, while pulmonary
fibrosis, hepatic fibrosis, hepatic cirrhosis, and systemic
sclerosis belong to the latter (11). We thought that, to
decide the direction of treatment for keloids, it was essential
to determine whether keloids belong to the former group or
the latter one. The activity of MMPs is regulated at the
three levels; transcription, zymogen activation, and inhibition
of proteolytic activity (11). As for the regulations at the
level of transcription, most MMPs are induced through activation
of nuclear AP-1 transcription factor (14-16). The AP-1-dependent
activation of inducible MMPs is potently inhibited by glucocorticoids
(17) and retinoids (18) at the transcriptional level. With
regard to the regulations at the level of extracellular zymogen
activation, latent precursors or zymogens of most MMPs are
proteolytically activated via exposure of the catalytic site
(19). As for the regulations at the level of inhibition of
proteolytic activity, non-specific inhibitors, such as α2-macroglobulin
and α1-antiprotease, as well as specific inhibitors, tissue
inhibitors of metalloproteinases (TIMPs), are responsible
for the inhibition (20).
It was reported that MMP-1 and MMP-8 activities were upregulated
in photoaging skin by repeated exposure to ultraviolet irradiation
(21, 22). However, tretinoin (all-trans retinoic acid) suppressed
upregulated MMP-1 in photoaging skin at the level of transcription,
probably via anti-AP-1 effects (23).
The activity of MMPs is also intimately correlated with the
invasive or metastatic ability of malignant tumor cells (24,
25). Especially for skin malignancies, degradation of ECM
is the first step to local invasion and metastasis. Thus,
basic and clinical studies have been performed with the aim
of chemoprevention of ECM degradation in malignant melanoma,
basal cell carcinoma, and squamous cell carcinoma (26-28),
as well as chemoprevention of cell growth. Retinoids are
the subject of increasing interest as an effective means
to control upregulated MMPs activity of malignant tumor cells
and inhibit the advancement of tumors (27). It has been reported
that retinoids suppress MMP-1 and MMP-8 activity in these
malignant tumor cells in vitro (28).
Thus, we hypothesized that degradation of ECM may be upregulated
during the expansion of keloids into circumferential skin,
and that high metabolic activity of keloid tissues (29) may
be due to increased MMPs activity, which may contribute to
continuous itching and/or tenderness associated with chronic
inflammation seen in keloids (30). Based on these hypotheses,
we examined differences in expressions of MMP-1, MMP-8, and
MMP-13 between keloid-derived fibroblasts and normal dermal
fibroblasts. Since retinoids are potent inhibitors of MMPs
in the treatment of photoaged skin and cancers as described
above, we also examined whether or not tretinoin affects
MMPs expressions of keloid derived fibroblasts.
Materials and Methods
Clinical Specimens
A total of 12 specimens of keloid (keloid group), diagnosed
on the basis of their clinical appearance, anatomic location,
etc., were excised at the Department of Plastic and Reconstructive
Surgery, the University of Tokyo Hospital. As a control group,
a total of 12 normal skin samples, matched to the site of
predilection for keloids (scapular area, shoulder, and upper
arm), were also excised during the plastic surgery.
Part of each tissue sample was used to establish a primary
cell culture, and the rest was used for histopathologic diagnosis.
All keloid samples displayed the histopathology diagnostic
for keloids. No hypertrophic scar was included in the materials.
The clinical data of the keloid group and the control group
are shown in Table 1. No significant difference in age between
the two groups was observed (unpaired Student's t-test; P=0.4907).
All the biopsies were taken in accordance with the Declaration
of Helsinki.
Primary Dermal Fibroblast
Cultures
The primary dermal fibroblast cultures from the keloids (n=12)
and control skin samples (n=12) were established by explant
method (31). For primary culture of keloid fibroblasts, marginal
portions of keloid lesions were used. Briefly, after removal
of the reticular layer of the dermis and epidermis from total
skin samples, the surface side of the papillary layer was
attatched to the culture dish, then the culture medium was
added and a cell culture was started (37?C, CO2 5%). Subculture
was performed 2 weeks after primary culture, when cell culture
reached to 60-70% confluence. Human fibroblasts were isolated
from the same skin specimens for explant after they were
separated from the epithelium, and grown in FGM (Fibroblast
growth medium), which consists of Dulbecco's modified Eagle's
medium (DMEM), 0.6 mg/ml glutamine, and 10% fetal calf serum
(FCS).Since the primary culture of dermal fibroblasts contained
a small amount of keratinocytes, the passages 3 to 5 were
used for the experiment.
Measurement of MMPs mRNA
expression by real-time PCR.
Real-time reverse transcriptase polymerase chain reaction
(Real-time PCR) assays (32) on the basis of SYBR Green Chemistry
(33, 34) were performed with ABI PRISMR 7700 Sequence Detection
System (PE Biosystems, Foster City, CA) to quantify the MMP-1,
MMP-8, and MMP-13 mRNA expressions.
The fibroblasts of the keloid group and normal group were
seeded at the density of 5×106 cells on a 100 mm Petri dish
in 10 ml of culture medium. Forty-eight hours after seeding,
the culture medium of each dish was changed to the medium
containing 1 μM tretinoin. Total RNA was obtained with RNeasyR
Mini Kit (QIAGEN, Hilden, Germany) as described before (35),
at 0, 6, 12, 24, and 48 hours after the medium change. In
order to eliminate any residual genomic DNA, RNase-Free DNase
Set (QIAGEN, Hilden, Germany) was also applied. The concentration
of each RNA sample was measured with Spectrophotometer V-530
UV/VIS (JASCO, Tokyo, Japan).
A reverse transcriptase reaction was performed using RNA
PCR Kit (AMV) Ver.2.1 (TaKaRa, Tokyo, Japan). Five micro-gram
of total RNA in a 100 μl of reaction mixture (final concentrations:
5 mM MgCl2, 1 mM dNTP Mixture, 1 U/μl RNase Inhibitor, 0.125
μM Oligo dT-Adaptor Primer, 10mM Tris-HCl, 50 mM KCl, pH
8.3) containing 25 U of AMV Reverse Transcriptase XL, was
incubated at 42 ?C for 30 minutes, followed by inactivation
of the enzyme at 99 ?C for 5 minutes with Program Temp Control
System PC-700 (ASTEC, Fukuoka, Japan). The control reaction
was performed simultaneously with an otherwise identical
reaction, but without reverse transcriptase.
Real-time PCR was performed on ABI PRISM 96-Well Optical
Reaction Plates (PE Biosystems, Foster City, CA). Sequences
of each oligonucleotide primers are shown in Table 2. All
PCR reaction mixtures contained 25 μl of TaqMan SYBRR Green
PCR Master Mix (2×) (PE Biosystems, Foster City, CA), 0.25
μl of forward primer (10 pmol/μl), 0.25 μl of reverse primer
(10 pmol/μl), 4 μl of each diluted sample, 20.5 μl of DDW
per well. PCR amplification of the identical sample was performed
with both specific primer pairs of the target MMP gene and
human glyceraldehydes-3-phosphate dehydrogenase (GAPDH) gene
on the same reaction plate. The PCR reaction was comprised
of 40 cycles, consisting of denaturing at 95 ?C (15 sec.),
then annealing/extension at 60 ?C (1 min.). In order to eliminate
the possibility of contamination of genomic DNA during extraction
of total RNA, the RNA extract before reverse transcription
was amplified in the same way as the control, and no amplification
was detected.
Measurement of Secreted
MMPs Protein by Enzyme-Linked Immunosorbent Assay
(ELISA)
The fibroblasts derived from keloid tissue or normal skin
were seeded at the density of 5×106 cells on a 100 mm Petri
dish in 10 ml of culture medium as described above. The culture
medium of each dish of the experimental group was changed
to a medium containing 1 μM tretinoin (containing 10 μl of
ethanol as a vehicle), or that containing only 10 μl vehicle,
at 48 hours after seeding. Before assay, 2 ml of each culture
supernatant was concentrated by freeze-drying using a Freeze
Dryer FRD-mini (Asahi Technoglass, Tokyo, Japan). Freeze-dried
supernatants were dissolved in the assay buffer for the ELISA
system (0.03 M H3PO4, 0.1M NaCl 1 % bovine serum albumin,
0.01 M EDTA). For MMP-1 assay, 10× concentrated samples were
prepared, and for MMP-8 and MMP-13, 20× concentrated samples
were prepared.
The culture supernatant of each dish was collected 96 hours
after the medium change. BIOTRAK ELISA MMP-1, MMP-8, and
MMP-13 System (Amersham Pharmacia Biotech, Buckinghamshire,
U.K.) was used for measurement of MMP-1, MMP-8, and MMP-13
protein levels in each culture supernatant, respectively.
Standard and concentrated samples (10×) were incubated in
microtiter wells precoated with a primary mouse anti-human
MMP-1 monoclonal antibody followed by a secondary rabbit
anti-human MMP-1 polyclonal antibody. The resulting antigen-antibody
complex was detected using horseradish peroxidase (HRP)-labeled
donkey anti-rabbit IgG, and the conjugate was quantified
by a colorimetric reaction with 3,3',5,5'-tetramethylbenzidine
(TMB) substrate. After stopping the reaction with 100 μl
of 1 M sulphilic acid, the resultant color was read at 450
nm with Microplate Reader Model 550 (Bio-Rad Laboratories,
Hercules, CA). All samples were assayed in duplicate, and
the concentration of the target protein in each sample was
determined by interpolation from the standard curve.
Statistical Analysis
All data are presented as mean ± standard error. The data
were statistically analyzed using Student's t-test. Differences
in the keloid group and in the normal group were tested
using a paired t-test. Differences between the keloid
group and the control group were tested using an unpaired
t-test. A value of p<0.05 was considered significant.
Results
MMPs mRNA expressions in keloid-derived fibroblasts and normal-skin-derived
fibroblasts.
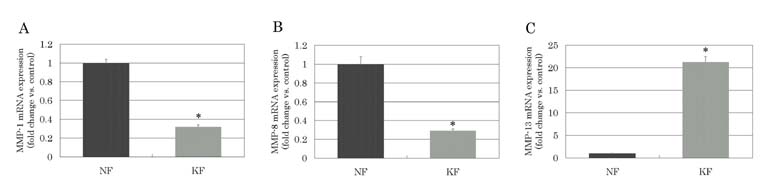
MMP-1, MMP-8, and MMP-13 mRNA expressions in the keloid group
and the control group were measured by real-time PCR system,
and the results are demonstrated in Fig. 1.
The normalized MMP-1 mRNA expression (MMP-1/GAPDH) was significantly
downregulated in keloid-derived fibroblasts compared to normal
fibroblasts (p=0.0001), and the fold change versus the average
of the control group was 0.32 ± 0.02 (mean ± standard error).
Similarly, the normalized MMP-8 mRNA expression was significantly
downregulated in keloid-derived fibloblasts (p=0.0120), and
the fold change versus the average of the control group was
0.29 ± 0.02. However, the normalized MMP-13 mRNA expression
was significantly elevated in keloid-derived fibroblasts
(p<0.0001), and the fold change versus the average of
the control group was 21.21 ± 1.24.
Effects of tretinoin on
MMPs mRNA expressions in keloid-derived fibroblasts
and normal-skin-derived fibroblasts.
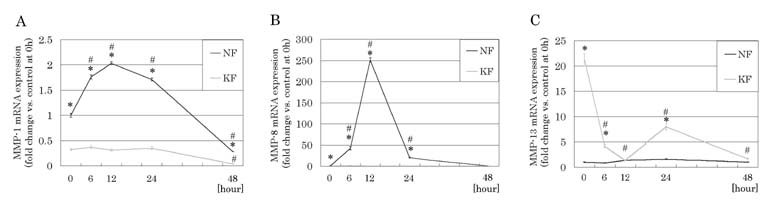
Effects of tretinoin on MMP-1, MMP-8, and MMP-13 mRNA expressions
over time were also examined by real-time PCR system, and
the results are shown in Fig. 2.
MMP-1 mRNA expression in the control group was significantly
upregulated with the peak at 12 hours after addition of tretinoin
(2.03 ± 0.03) (p<0.0001), whereas no significant change
was observed in the keloid group within 24 hours after the
addition of tretinoin. MMP-8 mRNA expression in the control
group was significantly upregulated with the peak at 12 hours
(250.80 ± 4.98) (p<0.0001), while no significant change
was observed in the keloid-derived fibroblasts after treatment
with tretinoin. In contrast, remarkably elevated MMP-13 mRNA
expression in the keloid group was significantly suppressed
with the peak at 12 hours by tretinoin (1.29 ± 0.04) (p=0.0003).
MMP-13 mRNA expression in the control group was not significantly
changed by treatment with tretinoin.
MMPs protein levels in
the culture supernatants and effects of tretinoin
on them
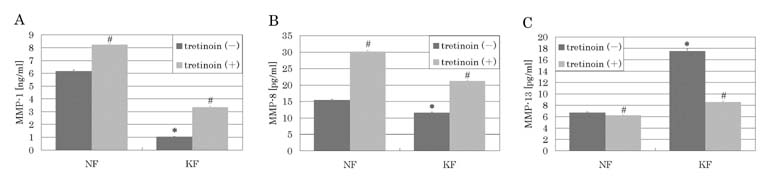
MMP-1, MMP-8, and MMP-13 protein levels in the culture supernatants
in the keloid group and the control group, and effects of
tretinoin on them were examined by ELISA. The results are
shown in Fig. 3.
MMP-1 protein expression was significantly lower in the keloid
group (1.04 ± 0.03 ng/ml) than in the control group (6.16
± 0.10 ng/ml) (p<0.0001). Similarly, the MMP-8 protein
level was significantly lower in the keloid group (11.54
± 0.24 pg/ml) than in the control group (15.36 ± 0.29 pg/ml)
(p=0.0043). However, the MMP-13 protein level was significantly
elevated in the keloid group (17.53 ± 0.33 pg/ml) in contrast
with the control group (6.71 ± 0.10 pg/ml) (p<0.0001).
In both the keloid group and the control group, the MMP-1
protein level was significantly elevated (3.35 ± 0.07 ng/ml,
8.22 ± 0.09 ng/ml) (p<0.0001, p=0.0019) by tretinoin
treatment for 96 hours. Additionally, both in the keloid
group and the control group, MMP-8 protein level was significantly
elevated (21.21 ± 0.22 pg/ml, 30.13 ± 0.37 pg/ml) (p<0.0001,
p<0.0001) by 96 hours' treatment with tretinoin. However,
the remarkably elevated MMP-13 protein level in the keloid
group was significantly decreased after treatment with tretinoin
for 96 hours (8.56 ± 0.20 pg/ml) (p<0.0001). The MMP-13
protein level in the control group was modestly suppressed
by tretinoin (6.23 ± 0.08 pg/ml) (p=0.0415).
Discussion
MMP-1, MMP-8, and MMP-13 all degrade type I and type III
collagen. Among the three MMPs, MMP-1 and MMP-8 most effectively
degrade type III and type I collagen, respectively. The decrease
in MMP-1 and MMP-8 may partly contribute to the accumulation
of type I and type III collagen in keloid tissues, and this
mechanism may be modulated by molecular interaction with
MMP-13.
MMP-13 is an abnormal collagenase subtype that has been found
in the bottom of chronic ulcers, where angiogenesis and fibrosis
occur (36). On the other hand, MMP-1 and MMP-8 are considered
to be "normal" collagenase subtypes that appear
in normal wound healing process (12, 37). Before the discovery
of MMP-13, reports had shown rather conflicting results concerning
to the collagenase activity to degrade type I or total collagen
in keloid tissues: some reports had shown normal (8), or
decreased (9), and others showed increased (10) activity
of collagenase. These variable results may be partly due
to different portions of keloid tissue, for example, a marginal
portion or a central portion. In our preliminary study, MMP-13
mRNA expression was found to be markedly higher in marginal
portions than central portions of keloid tissues (data not
shown). In the present study, comparison of MMPs expression
was performed using a marginal portion of each keloid sample.
Our study has demonstrated a significant increase in MMP-13
expression as well as a decrease in expressions of MMP-1
and MMP-8 in keloid-derived fibroblasts, both in mRNA and
protein levels. The remodeling of the surrounding matrix
by MMP-13 may interfere in normal degrading process of wound
healing in keloid tissues, and may initiate the negative
feedback mechanism to transcriptions of MMP-1 and MMP-8,
which act in the normal wound healing process. These mechanisms
could be related to chronic inflammation and infiltration
into circumferential normal skin seen in keloid tissues.
To correct the abnormal wound healing mechanism mentioned
above, we assumed that retinoids are potent additives, and
then investigated the influences of tretinoin on abnormal
MMP expressions of keloid tissues. The present study revealed
that addition of tretinoin to the culture media caused significant
downregulation of MMP-13 in keloid-derived fibroblasts at
both levels of mRNA and protein, and significant upregulation
of MMP-8 in normal dermal fibroblasts. Although mRNA expression
of MMP-1 was not clearly affected in the keloid-derived fibroblasts
by treatment of tretinoin, upregulation of MMP-1 and MMP-8,
and downregulation of MMP-13 at the protein level, may suggest
that tretinoin reverses the specific changes in the MMPs
expression profile of keloids. We also examined mRNA expressions
of four subtypes of TIMP (TIMP-1, -2, -3, and -4). All of
these subtypes were upregulated in keloid-derived fibroblats,
but we did not detect any significant changes after treatment
with tretinoin (data not shown).
A small number of past literatures reported effects of retinoids
on primary cultured human dermal fibroblasts. Daly et al.
(38) demonstrated that tretinoin significantly reduces collagen
production of human primary cultured fibroblasts. Abergel
et al. (39) reported that tretinoin and isotretinoin significantly
inhibit degradation activity of type I collagen fibers in
keloid-derived fibroblasts. On the other hand, in the field
of cancer cell study, degradation of type I and type IV collagen,
and invasion into collagen matrix was reported to be significantly
inhibited by retinoids (28). The results of our study and
those in the literature suggest that a remarkable inhibition
of degradation of type I collagen by tretinoin is presumably
due to a strong inhibition of MMP-13 expression by tretinoin,
which negates the upregulation of MMP-8.
Expressions of MMP-1 and MMP-13 are known to be induced at
transcriptional level by a variety of growth factors (14),
and these extracellular stimuli result in activation of nuclear
AP-1 trascription factor complex, which binds to the AP-1
cis-regulatory element in the promoter region of MMP gene
and potently activates transcription of the corresponding
MMP gene (15). This AP-1-dependent activation of inducible
MMPs is potently inhibited by glucocorticoids (17) and tretinoin
(18) at transcriptional level. The present results revealed
that MMP-13 was upregulated in keloid-derived fibloblasts
and this upregulation of MMP-13 was inhibited at the transcription
level by tretinoin, suggesting this upregulation of MMP-13
in keloids is induced via the AP-1 pathway. However, exactly
how tretinoin upregulates MMP-1 and MMP-8 in keloid-derived
fibroblasts, as well as in normal dermal fibroblasts, remains
unknown. Further investigations of the regulations are necessary
to clarify the mechanism.
In this study, it is suggested that MMPs are abnormally regulated
in keloid tissues as well as chronic ulcers, and that these
abnormal changes may be reversed by treatment with retinoids.
Tretinoin may improve chronic inflammation seen in keloids
and prevent expansion of keloid tissues into circumferential
normal skin.
Since 1999, we have been performing clinical trials with
tretinoin aqueous gel (0.1-0.4%) for treatment of keloids.
Our preliminary results demonstrated that topical application
of tretinoin on keloids has unique advantages. In most cases,
itching and/or tenderness of the lesions disappeared after
topical tretinoin (in preparation), although the volume-suppressing
effects on the fibrosis was quite modest. We assume that
effects of tretinoin on MMPs expression resulted in suppression
of chronic inflammation and prevention of growth and invasion
of keloid tissues. In considering limited clinical improvements
and side effects of existing techniques, the clinical use
of topical tretinoin looks promising. Thus, molecular mechanisms
of the regulation of MMPs deserve further investigation.
The results of this study may be helpful to develop more
chemically stable synthesized retinoids, which specifically
reverse abnormal expressions of MMPs and prevent cell growth
in keloids with minimal side effects.
Legends
Table 1. Profiles of skin
samples used in the experiment
control group number: 12
(normal skin) age: 17-51yrs. (32.8 ± 9.5 yrs.*)
sex: male: 6, female: 6
sites: scapular region: 9 ,
upper arm: 2, shoulder: 1
keloid group number: 12
age: 8-58 yrs. (29.0 ± 15.9 yrs.*)
sex: male: 5, female: 7
sites: scapular region: 4,
shoulder: 2, upper arm: 2,
chest: 2, forearm: 1, ear: 1
*: mean ± SD. No significant difference was observed between
the control group and the keloid group (p=0.4907).
Table 2. Oligonucleotid primers used in the real-time PCR
amplification of MMPs.
Gene Primer sequence*
Human MMP-1
ACGGATACCCCAAGGACATCT
CTCAGAAAGAGCAGCATCGATATG
Human MMP-8
ACCAAAGAGATCACGGTGACAA
TGAGCATCTCCTCCAATACCTTG
Human MMP-13
CCTGGAGCACTCATGTTTCCTAT
GACTGGATCCCTTGTACATCGTC
Human GAPDH#
GAAGGTGAAGGTCGGAGTC
GAAGATGGTGATGGGATTTC
*: All primer sequences
are written from 5' to 3'. For each primer pair,
the top sequence is sense and the bottom sequence
is antisense. #: GAPDH is human glyceraldehyde-3-phosphate
dehydrogenase and was used as a housekeeping gene.
|